
Concept de muscle
Les muscles forment le plus grand ensemble de tissus de l’organisme et représentent environ la moitié du poids du corps. Le tissus musculaire a une origine mésodermique et est constitué par des cellules allongées dénommées cellules musculaires, myocytes ou fibres musculaires, organisées en faisceaux enveloppés par du tissu conjonctif et spécialisées dans la contraction. Dans le muscle, la membrane cellulaire s’appelle le sarcolème et le cytoplasme est appelé sarcoplasme.
Il existe trois types de tissus musculaires classifiés de part leurs caractéristiques morphologiques et fonctionnelles (Fig. 1). On distingue les muscles striés squelettiques et cardiaque, qui présentent une striation transversale quand on les observe au microscope optique, et les muscles lisses (sans striation). D’un autre côté, on parle de muscles volontaires (squelettiques) et involontaires (lisses et cardiaque) selon qu’ils soient innervés respectivement par le système nerveux somatique ou par le système nerveux autonome.
Quelques propriétés sont partagées par les trois types de tissus musculaires: le système contractile est constitué par des filaments fins que glissent entre des filaments épais et la source d’énergie utilisée est l’adénosine triphosphate (ATP). Cependant, la structure et l’organisation des fibres ainsi que le mode d’excitation sont différentes dans les trois cas.

Fig.1. Représentation des différents types de muscles (Illustration par Anabela Fernandes, 2015)
- Muscle squelettique
Les cellules du muscle squelettique sont longues et cylindriques. Elles présentent plusieurs noyaux localisés à la périphérie proche de la membrane cellulaire (Fig. 1a). Le sarcolème est entouré par une lame basale et des cellules satellites. Celui-ci présente des invaginations profondes, appelées tubules transverses ou tubules T, impliquées dans la rapide dépolarisation de la membrane. Chaque fibre, caractérisée par le fait de contenir plusieurs myofibrilles, est entourée par une délicate couche de fibres réticuleuses, appelée endomysium, dans laquelle il existe un vaste réseau de capillaires sanguins et de nerfs. Les fibres musculaires se rassemblent en faisceaux entourés par une gaine fibro-élastique, le périmysium. Un ensemble de faisceaux musculaires forme ainsi un muscle entouré, à son tour, par une dense couche de tissus conjonctif appelée épimysium.
La plupart des muscles squelettiques, chez l’être humain, est formé par une combinaison des différents types de fibres musculaires. Les trois principales catégories de fibres sont:
– Les fibres du type I: Fibres rouges aérobiques de contraction lente et résistante à la fatigue dont le processus métabolique préférentiel est la phosphorylation oxydative;
– Les fibres du type IIa: Fibres intermédiaires, fibres de contraction rapide et modérément résistantes à la fatigue, avec une grande activité glycolytique et oxydative;
– Les fibres du type IIb: Fibres blanches anaérobiques de contraction rapide, assez sensible à la fatigue, qui utilisent de préférence la voie glycolytique.
La myofibrille, élément contractile spécialisé de la fibre musculaire, est composée par des éléments du cytosquelette disposés de forme régulière: les filaments épais et fins. L’unité fonctionnelle de ce système est le sarcomère (Fig. 2).
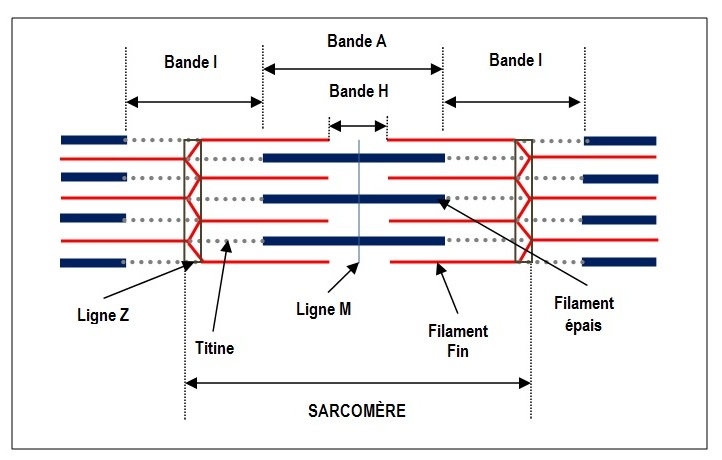
Fig. 2. Représentation schématique d’un sarcomère relâché (Illustration par Anabela Fernandes, 2015)
Les myofibrilles présentent une alternance de bandes sombres (bande A) et de bandes claires (bande I). Ces bandes se trouvent alignées donnant un aspect strié au muscle squelettique. Les bandes A, représentées sur la figure 2, sont constituées par des filaments épais entiers et par la superposition d’une partie des filaments fins. D’autre part, les bandes I sont constituées uniquement de filaments fins.
Les filaments épais sont composés de plusieurs centaines de protéines de myosine et les filaments fins sont constitués par trois protéines, l’actine (constituant le plus abondant), la troponine et la tropomyosine. Chaque molécule d’actine possède un site de liaison pour les ponts d’union de myosine.
L’élasticité du muscle et la stabilité des filaments fins et épais sont assurées par la présence des protéines de la ligne M et d’une protéine géante, la titine. Cette protéine, la plus grande de l’organisme (environ 30.000 acides aminés), est liée aux filaments de myosine et s’étend de la ligne Z à la ligne M.
Mécanisme de contraction musculaire
La stimulation du muscle squelettique est due à la libération d’un neurotransmetteur, l’acétylcholine, dans la jonction neuromusculaire (jonction entre la partie terminale d’un neurone moteur et la plaque motrice de la fibre musculaire). La liaison de l’acétylcholine aux récepteurs de la plaque motrice mène à la propagation d’un potentiel d’action (PA), à la superficie du sarcolème, qui pénètre à l’intérieur de la fibre à travers des tubules T.
Le réticulum sarcoplasmique forme un réseau discontinu de tubules fins qui englobe les myofibrilles. Les extrémités de chaque segment sont dilatées, formant des citernes latérales qui fonctionnent comme des réservoirs de calcium (Ca2+). Quand le PA arrive aux tubules T, celui-ci déclenche l’ouverture des canaux de Ca2+. Le Ca2+ libéré dans le cytosol se lie à la troponine provocant le déplacement de la tropomyosine, éloignant celle-ci du site de liaison des ponts de myosine sur l’actine. Les ponts d’union exécutent alors un mouvement de force qui permet le glissement des filaments d’actine et de myosine. Plusieurs cycles de contraction se passent pendant que les sites de liaison seront exposés et le Ca2+ disponible.
Quand les niveaux de calcium cytosolique diminuent, le Ca2+ se retire de la troponine et la tropomyosine reprend sa position originelle faisant que le muscle se détende.
L’énergie mécanique nécessaire au processus de contraction, dans les trois types de tissus musculaires, résulte de la transformation de l’énergie provenant de l’hydrolyse de l’ATP. Dans le muscle squelettique, cette hydrolyse requiert la liaison de magnésium (Mg2+) sur l’ATP car le Mg2+ fonctionne comme un accélérateur de l’activité de l’ATPase de la myosine. Cette enzyme clive l’ATP en adénosine diphosphate (ADP) et phosphate inorganique (Pi) (réaction de l’hydrolyse de l’ATP). L’énergie chimique libérée est stockée dans les ponts d’union et transformée en énergie mécanique nécessaire à la liaison de la myosine sur l’actine et au déplacement des myofilaments. L’ADP et le Pi sont rapidement libérés par la myosine. L’ATPase est alors disponible pour la fixation d’une nouvelle molécule d’ATP, ce qui mène à la rupture de la liaison entre l’actine et la myosine, revenant au début du processus.
Quand l’activité musculaire est intense, la quantité d’ATP, provenant de la respiration cellulaire et présente dans la cellule musculaire, n’est pas suffisante. La principale source de réserve énergétique du muscle est alors la phosphocréatine. La créatine kinase transfère un groupe phosphate de la phosphocréatine vers l’ADP pour former de l’ATP et de la créatine. D’autre part, quand il y a un excès d’ATP, la réaction inverse est catalysée, transférant un groupe phosphate de l’ATP vers la créatine, formant à nouveau de la phosphocréatine. Par conséquent, la quantité d’ATP libre dans le muscle, pendant une contraction, ne s’altère pas beaucoup.
Curiosités:
– Chez l’homme, les fibres musculaires sont plus épaisses et les muscles sont plus développés que chez la femme. Cela est dû à la testostérone, une hormone stéroïde sécrétée en plus grande quantité chez l’homme et qui favorise la synthèse de l’actine et de la myosine.
– Après la mort, l’absence d’ATP disponible fait que les liaisons entre l’actine et la myosine persistent, empêchant les cycles de contraction-décontraction. Ce phénomène produit une rigidité cadavérique ou rigor mortis. La rigidité commence entre 3 et 4 heures aprés la mort et est compléte aprés 12 heures. La décontraction a lieu aprés environ 36 heures.
- Muscle lisse
Le tissu musculaire lisse est disposé en couches ou faisceaux sur les parois de l’intestin, des voies biliaires, de la vessie urinaire, de la voie respiratoire, de l’utérus, des vaisseaux sanguins, dans l’iris et le corps ciliaire, dans le muscle érecteur du poil, entre autres. Les cellules de ce tissu sont plus courtes que les cellules du muscle strié, elles sont fusiformes et possèdent un noyau central (Fig. 1b).
Les cellules du tissu musculaire lisse possèdent trois types de filaments: les filaments épais de myosine, plus longs que dans le muscle squelettique, les filaments fins d’actine et tropomyosine mais dépourvu de troponine et les filaments intermédiaires, composés par de la desmine et la vimentine. Dans le cas du muscle lisse, ces trois types de filaments ne s’organisent ni en myofibrilles ni en sarcomères. De ce fait, le muscle lisse ne présente pas de striations.
Les filaments intermédiaires s’insèrent dans le cytoplasme et dans la membrane plasmique à travers des structures appelées corps denses. D’autre part, la membrane possède une grande quantité de dépressions de petites dimensions, les cavéoles, qui actuent comme un système primitif de tubules T et sont responsables du transport, par pinocytose, des ions Ca2+.
La myosine du tissu musculaire lisse est une myosine du type II, formée par deux chaines lourdes et deux paires de chaines légères. Elle se trouve enroulée, excepté quand elle est combinée avec un radical phosphate ce qui mène au dépliage du filament.
Après un stimulus donné, les niveaux de Ca2+ cytoplasmique augmentent. Le Ca2+ provient du réticulum sarcoplasmique et il entre aussi à partir du liquide extracellulaire. En suite, il se lie à la calmoduline formant un complexe qui active la kinase de la myosine. Cette dernière phosphoryle la chaine légère de la myosine qui se déroule exposant le site de liaison de l’actine sur la tète de la myosine. La myosine peut alors se lier aux filaments d’actine provocant la contraction musculaire. Quand les niveaux de Ca2+ sont diminués, la chaine légère de la myosine est enzymatiquement déphosphorilée et le muscle se détend.
Les cellules du muscle lisse sont liées les une aux autres à travers des jonctions communicantes qui permettent la contraction synchronisée du muscle lisse. D’autre part, les cellules sont recouvertes par une lame basale et sont englobées par un réseau de fibres réticulaires qui maintient les cellules unies de telle forme que la contraction de quelques cellules se transforme en la contraction du muscle entier.
Finalement, la contraction des cellules du tissu musculaire lisse est stimulée par des signaux nerveux, par la stimulation hormonale, par l’étirement du muscle, entre autres.
- Muscle cardiaque
Ce muscle est spécifique du cœur et est responsable de l’éjection du sang à travers les oreillettes et les ventricules ainsi que vers les vaisseaux sanguins. Les cellules du muscle cardiaque ou cardiomyocytes possèdent un noyau central, elles sont cylindriques et ramifiées (Fig. 1c). Les cellules s’interconnectent pour former des réseaux complexes. Elles sont plus petites que celles du muscle strié, avec environ 15µm de diamètre et 80 à 100µm de longueur.
Le muscle cardiaque possède des propriétés structurelles et fonctionnelles en commun avec le muscle squelettique. Les fibres cardiaques sont entourées par une délicate couche de tissu conjonctif, équivalent à l’endomysium du muscle squelettique, qui contient un abondant réseau de capillaires sanguins. Comme le muscle squelettique, le muscle cardiaque est strié, cependant les différences suivantes sont observées: les tubules T et le réticulum sarcoplasmique ne sont pas aussi développés; les cardiomyocytes sont unis aux extrémités à travers des connexions intercellulaires spécialisées, les disques intercalaires, qui sont des structures à forte coloration qui présentent une organisation en échelle, avec une portion centrale qui contient des desmosomes, et une portion longitudinale où se localisent les jonctions communicantes (Fig. 1c). Cette caractéristique est exclusive au muscle cardiaque.
Cependant, comme dans le muscle lisse, la contraction du muscle cardiaque est involontaire et le Ca2+ provient du réticulum sarcoplasmique ainsi que du liquide extracellulaire.
References:
- Gartner, L.P. and Hiatt, J.L . (2012). Histologia Essencial. Rio de Janeiro: Elsevier Brasil. p132-151.
- kierszenbaum, A.L. and Tres L.L. (2012). Histologia e Biologia Celular – Uma introdução à patologia. 3rd ed. Rio de Janeiro: Elsevier Brasil. p169-171.
- Sherwood, L. (2006). Physiologie musculaire. In: De boeck and Larcier. Physiologie humaine: A Human Perspective. Bruxelles: De boeck. p203-235.
- Widmaier, E.P. et al. (2003). Human Physiology – The mechanisms of body function. 9th ed. Boston: McGraw-Hill. p376; 450-454.


{kind=link}