
Concept of Muscle
Muscles are the largest set of body tissues and represent about half the weight of the body. Muscle tissue has a mesodermal origin and is composed of elongated cells, called muscle cells, myocytes or muscle fibres, which are arranged in bundles covered by connective tissue and are specialized in contraction. In the muscle, the cell membrane is called sarcolemma and the cytoplasm is called sarcoplasm.
There are three types of muscle tissue classified according to their morphological and functional features (Fig. 1). Muscles are classified as follow: skeletal striated and cardiac muscles, which have a transverse striation when observed by optical microscopy, and smooth muscles (not striated). On the other hand, they are referred as voluntary muscles (skeletal) and involuntary muscles (smooth and cardiac) whether they are enervated overwhelmed by the somatic nervous system or by the autonomic nervous system, respectively.
Some properties are shared by all three types of muscle tissue: The contractile system consists of thin filaments that slide between thick filaments and the source of energy used by the all three is the adenosine triphosphate (ATP). However, the structure and organization of the fibres and the excitement are different in all three cases.
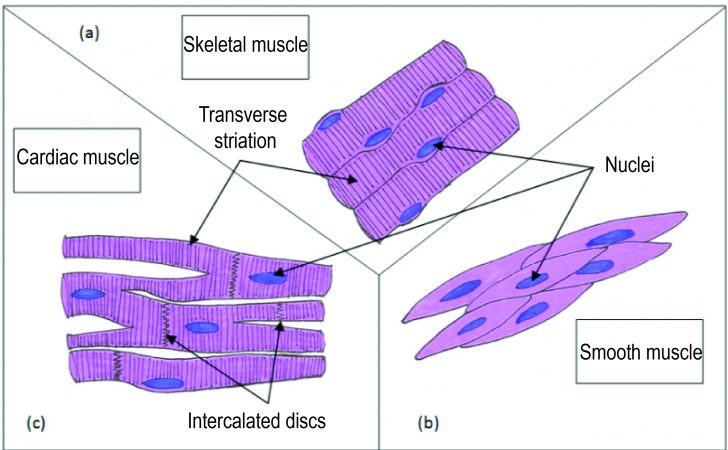
Fig. 1. Representation of the different types of muscles (Illustration by Anabela Fernandes, 2015)
Skeletal muscle
Skeletal muscle cells are long and cylindrical. It presents multiple nuclei located on the periphery close to the cell membrane (Fig. 1a). The sarcolemma is enveloped by a basal lamina and satellite cells. It has deep invaginations, called transverse tubules or T-tubules, involved in the rapid depolarization of the membrane. Each fibre, which has multiple myofibrils, is surrounded by a delicate layer of reticular fibres, called endomysium, in which there is an extensive network of blood capillaries and nerves. The muscle fibres join together into bundles or fascicles surrounded by a fibro-elastic sheath, the perimysium. A set of muscle bundles form a muscle surrounded, in turn, by a dense layer of connective tissue designated epimysium.
Most skeletal muscles in the human being, is formed by a combination of different types of muscle fibres. The three main categories of fibres are:
-Type I fibres: Aerobic red fibres with a slow contraction time and resistant to fatigue whose preferential metabolic process is the oxidative phosphorylation;
-Type Ia fibres: Intermediate fibres with a fast contraction time and moderately resistant to fatigue, with high glycoltic and oxidative activities;
-Type Ib fibres: Anaerobic white fibres with a fast contraction time, very sensitive to fatigue and which use preferably the glycol tic pathway.
The myofibril, specialized contractile element of the muscle fibre, is composed of cytoskeleton elements arranged on a regular basis: the thick and thin filaments. The functional unit of this system is the macromere (Fig. 2).
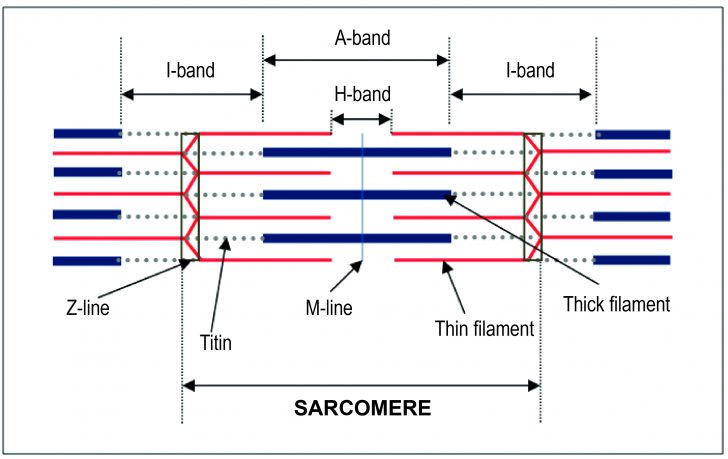
Fig. 2. Schematic representation of a sarcomere in relaxed state (Illustration by Anabela Fernandes, 2015)
The myofibrils present an alternation of dark bands (A-band) and light bands (I-band). These bands are aligned giving a striated appearance to the skeletal muscle. A-bands, as represented in figure 2, consist of entire thick filaments and overlaps of parts of thin filaments. In turn, I-bands consist of only thin filaments.
Thick filaments are composed of several hundred myosin proteins and thin filaments are composed of three proteins, actin (most abundant component), troponin and tropomyosin. Each actin molecule has a binding site for myosin cross-bridges.
The muscle elasticity and the stability of thin and thick filaments are guaranteed through the presence of M-line proteins and a giant elastic protein, called titin. This protein, the largest of the body (with about 30,000 amino acids), is linked to the myosin filaments and extends from the Z-line to the M-line.
Mechanism of muscle contraction
The stimulation of skeletal muscle is due to the release of a neurotransmitter, the acetylcholine, in the neuromuscular junction (junction between the end part of a motor neuron and the motor end plate of a muscle fibre). The binding of acetylcholine to receptors in the motor end plate leads to the propagation of an action potential (AP) on the surface of the sarcolemma and penetrates within the fibre through the T-tubules.
The sarcoplasmic reticulum forms a discontinuous network of thin tubules that enclose the myofibrils. The ends of each segment are dilated, forming lateral cisternae that act as reservoirs of calcium (Ca2+). When the AP reaches T-tubules, this triggers the opening of Ca2+ channels. The Ca2+ released in the cytosol binds to troponin causing displacement of the tropomyosin, shifted it away from the binding site of the myosin cross-bridges to the actin. The cross-bridges perform then a movement that allows the sliding of actin and myosin filaments. Several cycles of contraction happen while the binding sites are exposed and the Ca2+ is available.
When cytosolic calcium levels decrease, the Ca2+ turns off the troponin and tropomyosin goes back to its original position resulting in the muscle relaxation.
The mechanical energy necessary to the process of contraction, in all three types of muscle tissue, is the result of the transformation of energy from ATP hydrolysis. In skeletal muscle, this hydrolysis requires the binding of magnesium (Mg2+) to ATP as the Mg2+ works as an accelerant of the myosin ATPase activity. This enzyme cleaves the ATP into adenosine diphosphate (ADP) and inorganic phosphate (Pi) (ATP hydrolysis reaction). The chemical energy released is stored in the cross-bridges and transformed into the mechanical energy needed for the connection of the myosin on the actin and for the sliding of the myofilaments. ADP and Pi are released quickly by the myosin. ATPase becomes then available for the connection of a new ATP molecule, which leads to the break of the link between the actin and the myosin, returning to the beginning of the process.
When muscle activity is intense, the amount of ATP, from cellular respiration and present in the muscle cell, is not enough. The main source of muscle reserve energy is then the creatine phosphate. Creatine kinase transfers a phosphate group from creatine phosphate to ADP to form ATP and creatine. On the other hand, when there is an excess of ATP, the reverse reaction is catalyzed by transferring a phosphate group from ATP to creatine, forming the creatine phosphate. Consequently, the amount of free ATP in the muscle, during a contraction, does not change much.
Trivia:
– The muscle fibres are thicker and the muscles are more developed in men than in women. This is due to testosterone, a steroid hormone secreted in much greater quantity in men and that favours the synthesis of actin and myosin.
– After death, in the absence of available ATP, the strong biding state between actin and myosin persist, preventing contraction-relaxation cycles. This phenomenon produces the cadaverous rigidity or rigor mortis. The rigidity begins between 3 to 4 hours after death and is complete after 12 hours. Relaxation takes place after about 36 hours.
Smooth muscle
Smooth muscle tissue is found in layers or bundles in the walls of the intestine, the bile duct, the urinary bladder, the respiratory tract, the uterus, the blood vessels, in the iris and the ciliary body, the hair erector muscle, among others. The cells of this tissue are shorter than the striated muscle cells, they are fusiform, and have a central nucleus (Fig. 1b).
Smooth muscle cells have three types of filaments: thick myosin filaments, longer than in skeletal muscle, thin actin and tropomyosin filaments, devoid of Troponin, and intermediate filaments, composed by desmin and vimentin. In the case of smooth muscle, these three types of filaments are organised neither in myofibrils nor in sarcomeres. As such, smooth muscle does not have striations.
The intermediate filaments are inserted in the cytoplasm and in the plasma membrane through structures called dense bodies. On the other hand, the membrane has a large amount of small depressions, called caveolae, that act as a primitive system of T-tubules and are responsible for the transportation, by pinocytosis, of Ca2+ ions.
The myosin of the smooth muscle tissue is a myosin type II, composed of two heavy chains and two light chains pairs. This myosin is tangled, except when combined with phosphate that leads to the unfolding of the filament.
After a given stimulus, cytoplasmic Ca2+ levels increase. The Ca2+ comes from the sarcoplasmic reticulum and also enters from the extracellular fluid. Then, it binds to calmodulin forming a complex that activates the myosin kinase. The latter phosphorylates the myosin light chain that unfolds to expose the actin binding site located on the myosin head. The myosin can then bind to actin filaments causing muscle contraction. When Ca2+ levels are reduced, the myosin light chain is enzymatically dephosphorylated and the muscle relaxes.
Smooth muscle cells are connected to each other through gap junctions that allow the synchronized contraction of the smooth muscle. On the other hand, the cells are coated by a basal lamina and are surrounded by a network of reticular fibres that keeps cells together in such a way that the contraction of just a few cells leads to the contraction of whole muscle
Finally, the contraction of smooth muscle cells is stimulated by nerve signals, hormonal stimulation, muscle stretch, among other things.
Cardiac muscle
This muscle is the heart muscle and is responsible for boosting the blood through the atria and the ventricles and into the blood vessels. The heart muscle cells or cardiomyocytes have a central nucleus; they are cylindrical and branched (Fig. 1 c). The cells are interconnected to form complex networks. They are smaller than striated muscle cells, with approximately 15 µm in diameter and 80 to 100 µm in length.
The heart muscle has properties of structure and function in common with skeletal muscle. Cardiac fibres are surrounded by a delicate connective tissue layer, equivalent to the skeletal muscle endomysium, which contains a rich network of blood capillaries. As in skeletal muscle, the heart muscle is striated, however the following differences are observed: T-tubules and the sarcoplasmic reticulum are not as well developed as in skeletal muscle; the cardiomyocytes are united by the ends through intercellular connections, intercalated discs, which are heavily stained and structures that present a step-like arrangement, with a transverse portion, containing desmosomes, and a longitudinal portion where are located the gap junctions (Fig. 1 c). This is a unique feature of the heart muscle.
However, as in smooth muscle, contraction of the cardiac muscle is involuntary and the Ca2+ comes from the sarcoplasmic reticulum and the extracellular fluid.
References:
- Gartner, L.P. and Hiatt, J.L . (2012). Histologia Essencial. Rio de Janeiro: Elsevier Brazil. p132-151.
- kierszenbaum, A.L. and Tres L.L. (2012). Histologia e Biologia Celular – Uma introdução à patologia. 3rd ed. Rio de Janeiro: Elsevier Brazil. p169-171.
- Sherwood, L. (2006). Physiologie musculaire. In: De boeck and Larcier. Physiologie humaine: A Human Perspective. Bruxelles: De boeck. p203-235.
- Widmaier, E.P. et al. (2003). Human Physiology – The mechanisms of body function. 9th ed. Boston: McGraw-Hill. p376; 450-454.


{kind=link}