
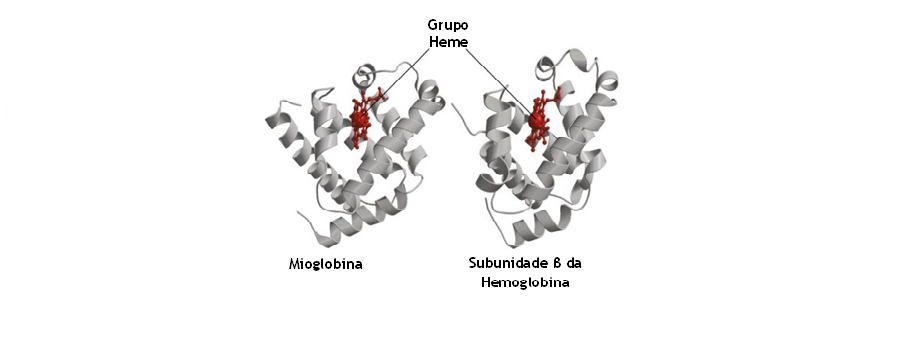
Estudo realizados através da técnica de Raio-X em diversas proteínas demonstrou uma indicação errônea de que as proteínas apresentavam estruturas rígidas e fixas. Na verdade, as proteínas são moléculas bastante flexíveis e com mobilidade estrutural. Por exemplo, estudos realizados a partir da técnica de Raios-X indicam que o grupo heme da hemoglobina e da mioglobina estão tão rodeados pelas proteínas que não há um caminho próprio para a molécula de O2 se aproximar ou “escapar” da zona de ligação. No entanto, sabe-se que a mioglobina e a hemoglobina conseguem promover facilmente a ligação e a libertação das moléculas de O2. Assim sendo, as estruturas tridimensionais da hemoglobina e da mioglobina evoluíram ao nível da sua flexibilidade por forma a facilitar a difusão de O2 da sua zona de ligação.
 Assim, as estruturas intermoleculares das proteínas têm sido classificadas em três grandes classes de acordo com a sua coerência:
Assim, as estruturas intermoleculares das proteínas têm sido classificadas em três grandes classes de acordo com a sua coerência:
- Flutuações atómicas, tais como as vibrações das ligações individuais, que têm períodos de tempo que variam entre 10-15 a 10-11 s e deslocamentos espaciais entre 0,01 a 1 Angstroms (Å).
- Movimentos coletivos, em que os grupos de átomos ligados covalentemente, (variam apenas no tamanho das cadeias laterais dos aminoácidos) se movem com variações entre 10-12 a 10-3 s e deslocamentos espaciais entre 0,01 e > 5 Å. Tais movimentos podem ocorrer com mais ou menos frequência em comparação com o seu período de tempo característico.
- Alterações conformacionais desencadeadas, em que os grupos de átomos (variam no tamanho das cadeias laterais dos aminoácidos) se movem em resposta a estímulos específicos, tais como a ligação de uma pequena molécula, por exemplo, a ligação da molécula de adenosina trifosfato (ATP) para GroEL (molécula que pertence à família das chaperoninas, importante no dobramento adequado de muitas proteínas). Assim sendo, as alterações conformacionais podem ocorrer ao longo de intervalos de tempo que variam entre 10-9 a 103 s com deslocamentos atómicos entre 0,5 e > 10 Å.
Todavia, o estudo da caracterização dos vários movimentos das proteínas é extremamente importante, uma vez que apresenta uma relevância a nível estrutural e funcional:
- As proteínas apresentam estruturas móveis – A análise cristalográfica de Raios-X é uma técnica poderosa para a análise do movimento das proteínas; esta técnica revela não só as posições médias dos átomos num cristal, mas também os seus deslocamentos médios nessas posições. A análise de Raios-X indica, por exemplo, que a mioglobina apresenta um núcleo rígido em torno do seu grupo heme e que as regiões mais periféricas da molécula apresentam um carácter mais móvel. Uma técnica “teórica” iniciada por Martin Karplus, denominada de simulação em dinâmica molecular, permitiu revelar a natureza dos movimentos atómicos em proteínas. Nesta técnica, os átomos de uma proteína de estrutura “conhecida” e ao seu meio envolvente (solvente aquoso) é lhes inicialmente atribuídos “movimentos aleatórios” com velocidades que são coletivamente características de uma temperatura escolhida.
- A mobilidade do núcleo da proteína é revelada através dos anéis aromáticos – A espectroscopia de ressonância magnética (RMN) pode determinar a mobilidade dos diferentes grupos das proteínas sob uma ampla gama de escalas de tempo. Consequentemente, a taxa em que um anel aromático numa proteína se desdobra é melhor caracterizada a partir da análise do seu espectro de RMN (como por exemplo, os anéis aromáticos de inversão não são detetados por cristalografia de raios-X, uma vez que esta técnica só revela a estrutura média de uma proteína).
- Modificações frequentes podem ser detetadas através da carga do átomo de hidrogénio – As alterações conformacionais que ocorrem ao longo de intervalos de tempo e que abrangem mais do que alguns segundos podem ser caracterizadas quimicamente através do estudo da interligação da proteína ao átomo de hidrogénio. Assim sendo, o interior das proteínas em grande parte devido às ligações com os átomos de hidrogénio não entram assim em contacto com o meio envolvente (solvente aquoso), o mesmo ocorre com os protões internos das proteínas. Por outro lado, quando ocorre a ligação dos protões internos das proteínas com o solvente, isto pode dever-se, a uma consequência do desdobramento transitório local que expõem estes protões “permutáveis” ao solvente aquoso.
Outros assuntos relacionados:
- Estabilidade das proteínas;
- Estrutura primária das proteínas;
- Estrutura secundaria da proteína;
- Estrutura terciária da proteína;
- Estrutura quaternária da proteína.
References:
- Anfinsen,C.B., Principles that govern the folding of protein chains, Science 181, 223–230 (1973).
- Henzler-Wildman, K. and Kern,D., Dynamic personalities of proteins, Nature 450, 964–972 (2007).
- Karplus, M. and McCammon,A., Molecular dynamics simulations of biomolecules, Nature Struct. Biol. 9, 646–651 (2002).
- Protein dynamics, Science 324, 197–215 (2009).




{kind=link}