
Conceito de Peroxissoma
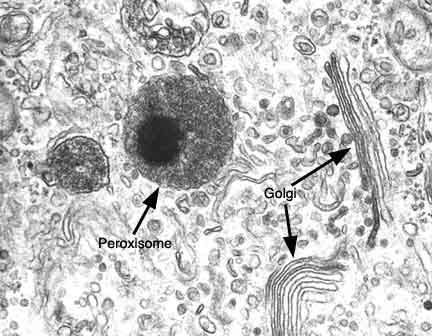
Os peroxissomas são organelos subcelulares ubíquos, que podem ser encontrados virtualmente em todas as células eucarióticas.
Apesar de estarem presentes em todos os tecidos, são encontrados em maior concentração no fígado e rins, devido ao seu papel na detoxificação de várias moléculas tóxicas presentes nestes órgãos.
Os peroxissomas são limitados por uma membrana simples de bicamada lipídica, embebida por proteínas de membrana peroxissomais. Apresentam no seu interior uma fina matriz granular envolvida em várias funções metabólicas, e que pode conter inclusões cristalinas de enzimas da matriz. Geralmente possuem um diâmetro entre 0,1µm e 1µm e apresentam uma morfologia esférica. Contudo, os peroxissomas apresentam uma grande plasticidade, podendo apresentar uma ampla variedade de formas, para além de possuírem a capacidade de modificar a sua morfologia em resposta a alterações fisiológicas no ambiente celular. Para além disso, possuem também a capacidade de adaptar o seu número, quantidade de enzimas e funções metabólicas conforme condições específicas.
Representação esquemática de um peroxissoma. De facult.une.edu.
Os peroxissomas são mais do que simples organelos metabólicos, são considerados organelos altamente dinâmicos, versáteis, metabolicamente ativos e que interagem funcional e morfologicamente com outros organelos, como as mitocôndrias, retículo endoplasmático (RE) e inclusões lipídicas. Por exemplo, os peroxissomas e as mitocôndrias mantêm uma estreita relação, que inclui cooperação metabólica, contribuição para a homeostasia celular e partilha de proteínas essenciais da maquinaria de divisão. Esta interação provavelmente influencia a funcionalidade de ambos os organelos.
Descoberta
Os peroxissomas foram descobertos em 1954 por Johannes Rhodin ao utilizar microscopia eletrónica em rim de ratinho. Inicialmente foram denominados por microbodies devido à sua morfologia, no entanto, em 1965, o citologista belga De Duve (Prémio Nobel, 1974) e os seus colegas isolaram com sucesso peroxissomas a partir de fígado de rato e descobriram uma co-localização de várias oxidases produtoras de peróxido de hidrogénio (H2O2), bem como catalase, enzima que degrada H2O2, na matriz destes organelos. Passou então a ser usado o termo funcional “peroxissomas” devido ao facto de estes organelos conterem uma ou mais enzimas que usam o oxigénio molecular para remover átomos de hidrogénio.
A vida do peroxissoma
Os peroxissomas possuem características únicas, que estão muitas vezes em desacordo com alguns dogmas existentes na biologia celular, designadamente em relação à sua biogénese e importação de proteínas, as quais diferem substancialmente de outros organelos. Os peroxissomas possuem a particularidade de as suas proteínas serem codificadas por genes nucleares, uma vez que não possuem moléculas de DNA, nem maquinaria associada à tradução de proteínas. Como tal, a maioria das proteínas peroxissomais é sintetizada no citosol em ribossomas livres e posteriormente, após a tradução, diretamente importada para os peroxissomas pré-existentes.
A biogénese dos peroxissomas envolve três principais etapas:
- formação da membrana peroxissomal;
- importação de proteínas para a matriz peroxissomal;
- proliferação do organelo.
Existem dois principais mecanismos propostos para a formação dos peroxissomas:
- Modelo “crescimento e divisão” – defende que os organelos se podem multiplicar a partir de peroxissomas pré-existentes, consistindo num processo de várias etapas que inclui elongação/crescimento dos peroxissomas, constrição e final fissão/divisão, dando origem a dois peroxissomas, geralmente esféricos que podem ser simétricos ou assimétricos;
- Modelo de formação de peroxissomas de novo – defende que os peroxissomas são capazes de formarem-se a partir do RE, ocorrendo a formação de um peroxissoma maduro por meio da fusão de duas vesículas pré-peroxissomais.

Vista esquemática da formação dinâmica do peroxissoma. De Koch, A., Schneider, G., Lüers, G. H., & Schrader, M. (2004). Peroxisome elongation and constriction but not fission can occur independently of dynamin-like protein 1. Journal of Cell Science, 117(Pt 17), 3995–4006.
Os peroxissomas possuem a capacidade de proliferar ou serem degradados em resposta a estímulos nutricionais ou do ambiente extracelular. Quando necessário, os peroxissomas são removidos por macroautofagia. Neste mecanismo de autofagia o peroxissoma é sequestrado dentro de autofagossomas, os quais posteriormente se fundem com os lisossomas/vacúolos, onde ocorre a digestão do organelo.
Funções
Os peroxissomas são considerados organelos multi-purpose, uma vez que possuem funções específicas que variam de acordo com o organismo e tipo celular, fase de desenvolvimento do organismo e condições ambientais.
Contribuem para diversas vias bioquímicas, nomeadamente uma ampla variedade de reações catabólicas e anabólicas, sendo essenciais para a saúde humana e desenvolvimento. Contribuem para a homeostasia celular dos ROS (espécies reativas de oxigénio), sinalização, desenvolvimento e envelhecimento. Recentemente, foi ainda demonstrado que os peroxissomas desempenham um importante papel na resposta imune celular, designadamente em parceria com as mitocôndrias na resposta inata a infecções virais.
As principais funções dos peroxissomas englobam o metabolismo do peróxido de hidrogénio e o dos lípidos, destacando-se nestes últimos a β-oxidação dos ácidos gordos.
A β-oxidação é o principal mecanismo de degradação dos ácidos gordos. Quando os ácidos gordos são mais complexos, como é o caso dos ácidos gordos de cadeia muito longa (VLCFA, Very-Long-Chain Fatty Acids), a sua degradação tem que ocorrer inicialmente nos peroxissomas. Este processo é semelhante ao que ocorre nas mitocôndrias, apesar de apresentarem algumas diferenças, como o facto de os peroxissomas não possuírem cadeia respiratória, e deste modo tornarem este processo num mecanismo de geração de calor. No entanto, os peroxissomas não possuem capacidade para completar o processo, sendo que o último produto derivado da via peroxissomal é posteriormente transportado para a mitocôndria, onde o processo de decomposição é finalizado.
A enzima catalase (CAT) está presente em praticamente todos os organismos aeróbios e é uma das enzimas mais importantes e presente em maiores quantidades nos peroxissomas. Esta peroxidase é importante para a regulação do stress oxidativo e inflamação, para além de proteger as células dos efeitos tóxicos do peróxido de hidrogénio (produto da β-oxidação e outras vias metabólicas que ocorrem nos peroxissomas e tóxico quando acumulado); para isto a catalase decompõe o peróxido de hidrogénio (H2O2) em água (H2O) e oxigénio (O2). A catalase também utiliza o H2O2 para oxigenar outros substratos, processo importante para a detoxificação de moléculas tóxicas. Outras enzimas envolvidas na produção e eliminação de ROS são a Superóxido Dismutase (SOD), as Peroxirredoxinas (Prx) e a Glutationa Peroxidase (GPx).
Os peroxissomas possuem também um importante papel noutros processos metabólicos, como por exemplo, a biossíntese de lípidos, α-oxidação de ácidos gordos, regulação da razão Acil-CoA/CoA, metabolismo de aminoácidos/proteínas e biossíntese de glicerol. Novas funções foram descobertas nos últimos anos, como a produção de feromonas e o metabolismo de poliaminas. Na tabela 1 é apresentada uma compilação de algumas funções dos peroxissomas.
Tabela 1 – Funções peroxissomais
| Funções clássicas |
| Metabolismo do peróxido de hidrogénio (catalase e oxidases que geram H2O2), metabolismo de ROS/NOS |
| Biossíntese de lípidos (fosfolípido éter/plasmalogénio, ácidos biliares, colesterol e dolicol, alongamento dos ácidos gordos) |
| β-oxidação de ácidos gordos (ácidos gordos de cadeia muito longa (VLCFA), ácidos dicarboxílicos, ácidos gordos de cadeia ramificada, ácidos gordos insaturados, metabolismo do ácido araquidónico e compostos xenobióticos) |
| α-oxidação de ácidos gordos (ácido fítico, compostos xenobióticos) |
| Ativação de ácidos gordos de cadeia longa/muito longa |
| Regulação da razão Acil-CoA /CoA |
| Metabolismo de proteínas/aminoácidos (biossíntese de cisteína e assimilação de enxofre, degradação do ácido aminado, metabolismo de L-lisina, degradação de poliamidas, proteases, transaminases) |
| Metabolismo de nicotinamida e nicotinato |
| Metabolismo de retinóide |
| Catabolismo das purinas |
| Biossíntese de glicerol |
| Biossíntese de ácido biliar |
| Detoxificação do glioxilato e metabolismo do dicarboxilato |
| Via hexose-monofosfato |
| Novas funções |
| Síntese de jasmonato (plantas) |
| Síntese de auxinas (plantas) |
| Produção de feromonas (animais) |
| Biossíntese de isoprenóides (plantas, animais) |
| Síntese de biotina (plantas, fungos) |
| Síntese de toxinas (fungos) |
| Metabolismo de poliaminas (plantas, animais e fungos) |
| Biossíntese de filoquinona (vitamina K1) (plantas) |
| Síntese de glicina betaína (plantas) |
| Defesa imune inata em infecções virais (animais) |
| Biossíntese de âncora GPI (animais) |
| Sinalização (H2O2) em neurónios do hipocampo (animais) |
Doenças peroxissomais
Existem muitas desordens associadas aos peroxissomas, e muitas vezes uma simples alteração num gene pode conduzir a uma severa doença, por vezes mesmo letal. As desordens peroxissomais são geralmente classificadas em dois grupos:
(1) Doenças da Biogénese Peroxissomal (DBPs) – grupo de desordens cerebrais de desenvolvimento com uma prevalência de 1:50000 e que geralmente conduzem à morte durante a infância. Estas doenças são causadas por mutações nos genes PEX que codificam peroxinas, proteínas necessárias para a biogénese dos peroxissomas e para a importação de proteínas da matriz e da membrana peroxissomal. Consequentemente, os peroxissomas podem estar completamente ausentes devido a um defeito na sua formação ou estarem presentes como compartimentos membranares vazios (“ghosts”) devido a defeitos na maquinaria de importação das enzimas peroxissomais. Este grupo de doenças inclui o espectro Zellweger que engloba o Síndrome de Zellweger (SZ), a Adrenoleucodistrofia Neonatal (ALDN) e a Doença de Refsum Infantil (DRI); e o espectro CDPR do tipo I que engloba a Condrodisplasia Rizomélica Punctata do tipo I (CDPR).
(2) Doenças peroxissomais com alterações funcionais numa única enzima – baseadas na ocorrência de mutações num único gene que vão consequentemente afetar a atividade e/ou localização da proteína peroxissomal correspondente, sendo que apenas uma função metabólica vai ser comprometida. Como tal este grupo de doenças está associado à acumulação de substratos tóxicos ou à escassez de produtos peroxissomais. Podem ser subdivididas em distintos subgrupos em função da via metabólica peroxissomal afetada: biossíntese de fosfolípido éter (plasmalogénio), β-oxidação de ácidos gordos, α-oxidação peroxissomal, detoxificação do glioxilato e metabolismo do H2O2.
References:
- Alberts, B., Johnson, A., Lewis, J., Raff, M., Roberts, K., & Walter, P. (2002). Peroxisomes. Garland Science.
- Islinger, M., & Schrader, M. (2011). Peroxisomes. Current Biology : CB, 21(19), R800–1.
- Masters, C. (1995). The Peroxisome: A Vital Organelle (p. 286). Cambridge University Press.
- Schrader, M., & Fahimi, H. D. (2008). The peroxisome: still a mysterious organelle. Histochemistry
and Cell Biology, 129(4), 421–40. - Islinger, M., Grille, S., Fahimi, H. D., & Schrader, M. (2012). The peroxisome: an update on
mysteries. Histochemistry and Cell Biology, 137(5), 547–74. - Wanders, R. J. a, & Waterham, H. R. (2006). Biochemistry of mammalian peroxisomes revisited.
Annual Review of Biochemistry, 75, 295–332. - Schrader, M., & Fahimi, H. D. (2006). Peroxisomes and oxidative stress. Biochimica et Biophysica
Acta, 1763(12), 1755–66. - Baker, A., & Paudyal, R. (2014). The life of the peroxisome: from birth to death. Current Opinion in
Plant Biology, 22, 39–47. - Van der Zand, A., Gent, J., Braakman, I., & Tabak, H. F. (2012). Biochemically distinct vesicles from
the endoplasmic reticulum fuse to form peroxisomes. Cell, 149(2), 397–409. - Veenhuis, M., & Klei, I. J. Van Der. (2014). De novo peroxisome biogenesis revisited, 1(4), 128–
130. - Hettema, E. H., & Motley, A. M. (2009). How peroxisomes multiply. Journal of Cell Science, 122(Pt
14), 2331–6. - Delille, H. K., Bonekamp, N. a, & Schrader, M. (2006). Peroxisomes and disease – an overview.
International Journal of Biomedical Science : IJBS, 2(4), 308–14. - Wanders, R. J. a. (2004). Metabolic and molecular basis of peroxisomal disorders: a review.
American Journal of Medical Genetics. Part A, 126A(4), 355–75. - Camões, F., Bonekamp, N. a., Delille, H. K., & Schrader, M. (2009). Organelle dynamics and
dysfunction: A closer link between peroxisomes and mitochondria. Journal of Inherited
Metabolic Disease, 32(2), 163–180. - Odendall, C., & Kagan, J. C. (2013). Peroxisomes and the antiviral responses of mammalian cells.



{kind=link}